The Fundamental Laws of Human Behavior
TENTH LECTURE
Max Meyer
Table of Contents | Next | Previous
Order of acquisition of the first four classes of habits. Control of the sense organs of the head. Direction and extent of the fixation movement of the eye. Improvement by experience of the fixation movement of the eye. A variation of response resulting from a close succession of nervous processes just as from a deflection of one by another. Coordination of the eyes. How an infant learns to face a sounding object. Control of the hands and arms. Learning to raise the arm in response to a visual object above the eyes. Loose co-ordination of eye and hand.
WE have considered the change of behavior, which we call learning, in its positive as well as in its negative aspect. We have recognized that learning and unlearning (inhibition) are only the positive and the negative aspect of the same nervous function. We have further convinced ourselves that the three classes of change of behavior, which seem to include all possible kinds, variation of response, sensory condensation, and motor condensation, are at bottom the same nervous function, which in its simplest form we have called variation of response. The above explanation of this nervous function, however, is applicable only in case one nervous process which influenced another, begins before the other has ceased, that is, in case of simultaneity. Learning as applied to successive nervous processes will be explained farther below. We shall
( 121) first make ourselves acquainted with the most fundamental reflex activities of childhood from which human learning develops. The study of this development will show us in what respects the explanations of human behavior given thus far are sufficient and in what respects they need to be supplemented.
Childhood is the very age of learning in human life. Great as the accomplishments of human beings may appear which are acquired in schools and colleges, they are as a matter of fact small when compared with those of the first six or seven years, before the individual is systematically trained by his teachers. It is of the utmost importance for the teacher to understand how learning proceeds before it is systematically directed, in order to avoid the forcing of the individual into an educational system which is unrelated—or even opposed—to the natural way in which man acquires his habits in early childhood. The first habits acquired by the baby are those connected with the use of the sense organs of the head. Then the baby learns to use his arms and hands. Toward the end of the first year or soon thereafter he learns to use his 'feet for walking and running, and toward the end of the second year to use his vocal organs for speech. Of course, we do not mean that each one of these classes of activities must become perfect before the next begins, but merely that they usually become conspicuous in this order. Besides, it is well to keep in mind that individual differences are exceedingly common, so that these four stages of development are not in every child equally obvious.
Of the sense organs of the head, the eyes are those whose efficient use depends most on proper motor adjustments, —much more so than the ears or the sense organs of the mouth. The chief reflex of the eye consists in a turning of
(122) the center of the eye, the "fovea," the area of most distinct vision., in the direction of the most effective object of visual stimulation, in a "movement of fixation" as we may call it. It is quite natural that this response at first is often rather inadequate in accuracy of direction and particularly of extent,—the center of the eye moves either beyond the point where it could receive the optical rays from the object in question or not far enough to reach this point. Gradually, however, the movement becomes more and more adequate. Yet it never becomes absolutely exact even in adult life. Even the grown person, in order to fixate an object, must, practically without exception, correct the first sweeping movement of fixation by smaller ones before a sufficiently accurate adjustment of the fovea is obtained. Figure 37 gives an example of a complex eye movement of this kind, the first sweep of the object's image being from S,, to Sb.

Two things need explanation. First: why the eye ball moves always in the direction of the stimulus. Secondly: why the eye ball stops at the moment when the fovea is approximately opposite the stimulus. The first is easily explained. The various sensory points of the retina are doubtless connected with the eye muscles by reflex arches in such a way that, when any point of the retina is especially stimulated, the excitation takes its path toward those muscles or groups of muscles which pull the eye in the
( 123) proper direction. In this respect no learning is necessary. The direction is determined reflexly. The second question is less easily answered. Why does the eye movement stop as soon as the fovea has approximately reached a point opposite the object? One feels inclined to think that (luring the fixation movement of the eye the object, suddenly striking with its rays the fovea, sets up there a particularly effective excitation which either results reflexly in a muscular contraction capable of fixing the eye in the position just reached, or stops the movement by inhibiting the nervous process which causes it. Such explanations could still be offered as recently as ten years ago. But the research of the most recent years has discredited them. For some reason or other sensory excitations received while the eye moves, fail to become effective and, therefore, can not in any way stop the movement. The extent of the sweeping movement of the eye must also be determined reflexly and in advance of the motion—probably by a strong excitation reaching the muscles which pull the fovea in the proper direction and a relatively weak excitation reaching the antagonistic muscles. If the inherited connections of every point of the retina with the various groups of muscles are such as to determine through their resistances a definite ratio of the two antagonistic muscular effects just mentioned, the eye must clearly stop at a definite point,—where the antagonistic motor effects balance each other. Figure 38, for clearness' sake, suggests how the nervous conductors might be arranged in order to bring about this result; but we do not, of course, assert that they are arranged just in this way. The figure contains only two motor points, ML and MR, representing, say, the muscles which pull the eye to the left and those which pull to the right. The sensory points represent a series from one side of the retina, at
(124)
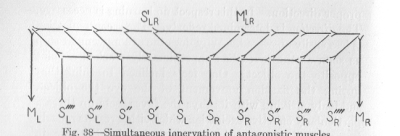
SL'''' (the right side, opposite the left edge of the field of vision) to the other side, at SR''''. It is evident that a nervous process starting, for example, from SL'', must go mainly to ML, traveling thus over the smallest number of neurons possible; but a considerable fraction of it must pass over S1LRM1LR to MR. A nervous process starting, for comparison, from S’L can send only a smaller part of its total flux to ML, since the resistance of the path from S’L to ML is greater than that of the path of fewer neurons from S’’L to ML: so a larger fraction of the flux takes the now shorter path over S1LRM1LR to MR. The farther we go to the sensory points at the right of the figure, the more we find MR favored in comparison with ML. Thus the eye must come to a standstill at different angles according to the sensory point excited and the corresponding ratio of the excitation of the antagonistic muscle groups.
This is simple enough thus far. However, this ratio does not seem to be established by inheritance with any great exactness, so that there is still left the problem which is of special interest to us in this study, namely, how we can explain the great improvement in the accuracy of the fixation movement which is to be observed during the first weeks and months of life,—how the child learns to execute these sweeping fixation movements with so much more accuracy in direction and extent after a few months
(125) of experience. The very purpose of our present study is, not to take learning as a phenomenon which is too common to need explanation, but to arouse our curiosity about it and to satisfy this by making the process of learning as plain as the running of a steam locomotive.
We may restrict our discussion to the extent of the movement stopping at, or before, or beyond a certain point, but always on the straight line passing through it. A movement along a line leaving the point to be fixated more or less at one side, is obviously simply the component, geometrically, of two movements, two muscular effects, and does not involve any additional principle of explanation. We shall, then, have to solve the following typical problem: If in earliest childhood, the fixation movement brings the image, let us say, from Sa, in Figure 37, to Sb, some distance beyond the fovea, why is it that after some experience the sweep is so much shorter that the image is brought from Sa to the nearest neighborhood of the fovea?
Figure 39 will aid us in understanding how the fixation movement of the eye can be improved by experience—in. understanding, at the same time, what the often mysterious word "experience" really signifies. Sa represents the sensory point where the image of the object first appears on the retina, Ma the motor point which signifies pulling the fovea in the direction of the optical image. Mb represents the motor point which signifies pulling the fovea in exactly the opposite direction. In order to bring about the proper movement of approximately the right extent, both Ma and Mb must be excited,—but Ma more strongly (if equally, there would be no movement at all), Mb only strongly enough to bring the eye ball after some time to a standstill, through the balancing of the opposite forces of muscular tension. The antagonistic muscles.
(126) in such a case, must act. upon the eye ball in the same way in which two rubber bands of different tension, attached to and pulling, say, a piece of cork in opposite directions, would act. We have assumed that the tension resulting from Mais relatively too great, so that the image, instead of falling upon the fovea, now falls at a point Sb, somewhat beyond the fovea. It is easy to understand from
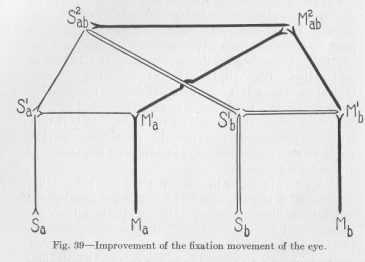
Figure 39 why stimulation of Sa should result in a stronger excitation at Ma and a weaker one at Mb. But why should, at a later recurrence of the stimulation of Sa, after the "experience" has become effective, the excitation at Mb be relatively greater, thus balancing the effect of Ma sooner and preventing the fovea from moving as far as the first time, stopping it nearer the correct point of fixation, the point F in Figure 37? The answer to this question is the solution of our problem.
(127)
In order to give the answer, we must consider the correcting movement. As soon as the eye stops after having swept too far, Sb is stimulated by the new optical impression, and—everything being exactly reversed—the fovea is pulled in the opposite direction. Let us see what neurons conduct the excitation whenever Sb is stimulated. They are, eight in number, marked in Figure 39 partly as broad, partly as double lines. A moment ago Sa was stimulated and the excitation took its path over some of these very neurons, namely, over those drawn in broad lines. The effect of that excitation, reducing the resistance of the neurons, lasts for some time before the negative susceptibility begins to restore the former resistance, as discussed previously and illustrated in Figure 33. Therefore, when now Sb is stimulated and the excitation coming from Sb reduces the resistance in all the neurons through which it passes, the resistance of the neurons drawn in broad lines not merely begins to be reduced, like the resistance of the neurons drawn in double lines, but is further reduced. In the total system of Figure 39, the neurons drawn in broad lines thus suffer a relatively greater reduction of their conductivity than all the others. It is easy to see that our problem is herewith solved, for when now at any time the retinal point Sa is again struck by an optical image, a greater fraction than originally of the excitation from Sa can pass over SabMab and MabMb into the non-corresponding point Mb, causing a stronger muscular tension at Mb and an earlier balancing of the antagonistic forces which bring about the sweeping movement of fixation.
To remove all doubt as to the correctness of this conclusion let us look again at the broad lines of Figure 39. The short route from Sa to Ma consists of three neurons, of which one is favored by the effect in question; that is, only
(128) one-third of the total length of the path. The longer route from S¢to Ma and the route from Sa to Mb, however, consist each of five neurons, of which three are favored by the effect in question; that is, as much as three fifths of the path. It is clear that the total result of this widening of one-third of the length of one path and of threefifths of the length of the other path is to the advantage of Mb, to the disadvantage of Ma. The ordinary superiority in intensity of response (to stimulation of Sa) of Ma over Mb is based on the greater conductivity of the reflex arch Sa S1a M1a Ma, which has now been relatively diminished. Any relatively greater reduction of the resistance of the higher center S2ab M1ab must favor the ordinarily quite weak response of the non-corresponding point Mb, to the disadvantage of the corresponding point Ma. The motor effect of Mbnow balances the effect of Ma a little earlier, brings about the stopping of the eye movement a little earlier. That is, the next fixation movement is more correct.
We see, then, that the improvement of any particular nervous function by "experience" of the individual is an altogether natural phenomenon,—no more mysterious, although different in kind, than the gradual improvement of the draft in a cold chimney after the fire has been burning for some time. A few fundamental laws which we assume as governing nervous activity are sufficient to explain this learning by experience, without any necessary reference to "conscious" experience. The present case is clearly a variation of response, for we have now a relatively strong motor effect where (at Mb in response to a stimulation of Sa) we had originally only a weak part of the total effect. But we notice the important fact that the variation has come about without a deflection of one nervous process by a simultaneous one taking
(129) place. We see, then, that the same variation which can be the result of a deflection, can also be the consequence of a succession of nervous processes, provided the law illustrated in Figure 33 is applicable, the law that any reduction in resistance remains for a short time (a few seconds at least) uninfluenced by the negative susceptibility.
We have explained how the improvement of the accuracy of the fixation movement comes about, which diminishes the need for movements of correction succeeding the first sweep. Simultaneously another improvement takes place,—that of the co-operation of the two eyes. During the first days of a baby's life it may not infrequently be observed that the eyes move independently of each other, that one moves to the right while the other stands still or moves even to the left. This is but natural since a striking object may not impress one eye as strongly as the other, indeed may be invisible to one eye if it happens to be near the right or left edge of the total field of vision. How, then, is the nervous system "educated" so that later both eyes always move together to the right or left, even when the object is, in advance of the movement, invisible to one eye?
In Figure 40, Ml and M, represent those motor points whose simultaneous excitations cause the simultaneous and similar movements of the two eyes. Whenever Sl and Sr, are stimulated simultaneously and with equal intensity, as it happens most frequently, the simultaneous excitations of the two motor points are self-evident. But why are, after a few months of life of the individual, the motor points simultaneously excited even when one of the sensory points is unstimulated? This is clearly a case of sensory condensation. It may be thought to come about in the following way.
( 130)
Whenever Sl is stimulated alone, by far the largest part of the excitation takes its path over the reflex arch directly to Ml. A smaller, though considerable, part, however, takes the indirect path over S2l M2l and even over S3lr M3lr , mostly in the direction of Ml, but to some extent also in other directions, for example, toward Mr .
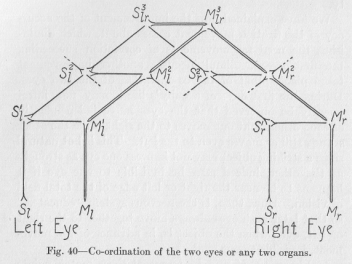
Whenever Sr is stimulated alone, by far the largest part of the excitation takes its path over the reflex arch directly to Mr. A smaller, though considerable, part, however, takes the indirect path over S2r M2r and even over S3lr M3lr mostly in the direction of Mr but to some extent also in other directions, for example, toward Ml. In either case, when Sl or Sr is stimulated alone, the relative resistance of the lower and higher neurons involved remains unchanged, in accordance with the law which we assumed in the preceding lecture. But when Sl and Sr are stimu-
( 131) -lated at the same time, those neurons which belong to the system of S, as well as to that of S, are favored over all the others with respect to the reduction of their resistance, since a double excitation and therefore a stronger flux passes through them. These neurons are drawn in Figure 40 in double lines. The relatively greater reduction of the resistance of these neurons may amount to but little in a single case of "experience." But it must become quite considerable in a few months, since simultaneous stimulation of both eyes by the same object is naturally a very common occurrence. When the resistance of the double drawn neurons has become much reduced, an ever increasing amount of the flux takes its path, instead of over a reflex arch, over the higher center S3lr M3lr, which forms the connecting bridge between the two eyes. Accordingly, an ever increasing part of the flux finds its way from M3lr. down into Mr, even when Sl happens to be stimulated alone, or into Ml when Sr, happens to be stimulated alone, until finally the response occurs regularly at both Ml and M, no matter whether both the sensory points are stimulated or only one of them. That is, the movements of the eyes have become perfectly co-ordinated.
The co-ordination of the eyes is an example of that kind of variation of the nervous path which we have called sensory condensation. Another example of sensory condensation which we have studied in detail is that of a child learning to execute, on the piano keyboard, two finger movements in response to only one note. The explanation in detail of the two examples is somewhat different owing to the fact that in the former case the sensory condensation results from a two-fold stimulation of unequal intensities, the one and the other sensory point being alternately more strongly excited, whereas in the present case both sensory points are usually stimulated with the same
( 132) intensity. We have thus seen that sensory condensation can result under various circumstances, provided only that the reflex Birches involved arc connected—and not too remotely—by higher nerve centers.
Next to the eye, the car is the sense organ of the head which is most interesting because of its motor adjustments. We are naturally inclined to assume that there is here a reflex similar to the turning reflex of the eye,—that, when a sound strikes one of the ears more strongly than the other, the face turns reflexly toward the source of sound. This would mean that, by inheritance, of the muscles turning the head those on either side are more closely connected with the one ear than with the other ear. The purpose of this head movement would be, not so much a better adjustment of the organ of hearing, as of the organ of sight, the eyes when facing the source of sound would be more likely to receive from it an effective visual stimulation. Such a reflex response, however, a turning of the face in the direction of a sound, does not seem to exist. When a child is five to six months old, this response is quite common, but during the first three months it does not occur. One may, of course, assume that the reflex does not mature until several months after birth, and support this assumption by the fact that some instincts and reflexes mature even years after birth. However, reflexes whose maturity is undoubtedly delayed take for example the sexual—are obviously delayed because their appearance directly after birth would be useless. But turning the head in the direction of a sound would not be entirely useless even in earliest infancy. We may therefore, as long as the question is left undecided by the physiologists and anatomists, regard it as most probable that the turning of the face in the direction of a sound is not a reflex, but a habit which is usually established three
(133) or four months after birth. Our problem, then, is to explain how the individual learns to respond thus to sounds which stimulate the two ears unequally.
There can hardly be any doubt that the habit of turning the face toward a source of sound is a variation of response —one of that kind in which the same response occurs, but another stimulus has been substituted for the original one. The original stimulus is visual. The fixation reflex of the eyes can be observed as functioning a few weeks after birth; and the reflex turning of the head can be seen to support and supplement the eye movement at the same early age,—in response to sight, for reflex movements of the head in response to tactual stimuli occur even a few days after birth, when the baby, held to the mother's breast, hunts, so to speak, for the nipple. Objects which appear in indirect vision and therefore call forth the fixation reflex, are frequently sources of sound, for example, the mother's face. That is, the baby's ears are stimulated at the same time as the eyes. If the nervous flux coming from the ear more strongly stimulated, and including, by deflection, most of the flux from the other ear, has no particular reflex outlet, it must take its path, other things being equal, mostly in the direction of any other strong nervous process existing at the same time and therefore attracting it. It must take its path in the direction of that motor point which in response to the visual stimulation turns the head to face the source of sound. There is no danger that the reverse might occur, that the nervous flux coming from the eye might be attracted by the flux from the ear, for we are working under the assumption that the flux from the ear has no low-resistance reflex outlet and, therefore, cannot attain any such great intensity as that from the eye. When the deflection from the ear to the head-turning muscles has
(134) occurred often enough and the resistance along this path has been effectively lowered, the habit which we intended to explain is established. The head then turns even in response to source of sound which may remain permanently invisible. That the habit is not established until several months after birth, is caused probably by the absence, at an earlier time, of connecting neurons which are capable of serving as a chain of conductors. It is a well established fact that the majority of the higher nerve centers capable of serving such remote connections are immature at birth and fairly mature only three months later.
The next large group of activities we proposed to study are those of the hands and arms. We have already seen how, from the reflex of a bending of the fingers in response to a stimulation of their inner surfaces, the delicate and skillful grasping of the older child and of the adult develops. We have also mentioned that the arm is reflexly stretched out in response to a stimulation of the eye. With respect to the arms, there are several further details which are interesting enough to be mentioned here.
With an adult, the raising of the hand in response to sight, as when we take a book from a shelf above our head (Figure 41), is a very common movement. During the
( 135) first months a similar response to sight, moving the hand in the direction from the feet to the head, does not occur. The movements like many other properties—of a human baby have often been compared with those of full-grown animals, for example, the apes; and, since the apes are climbers, one might conclude that the movement of Figure 41, which is common in climbing, must be frequently observed in babies. We see here, that, as a matter of fact, one has to be very critical in such comparisons. The apes are indeed climbing animals and, as such, are compelled to execute the arm movement in question quite frequently; but only in adult life, the baby apes do not climb any more than human babies, but cling to their mothers who carry them about. The comparison is therefore not really between babies and apes, but only between human babies and monkey babies. When the very young baby stretches out his arms, in response to a visual impression, it is always either at right angles to the longitudinal axis of the body or more downwards, toward the feet, never more upwards, toward the head. The same is probably true for the monkey baby.
How, then, does the baby learn to raise his hands in response to a visual stimulation? In order to answer this question, we must inquire whether there is during the first few months any reflex raising of the hands, and whether there is any possibility of this reflex being varied so that a visual stimulation takes the place of the stimulation of this reflex. There is a reflex of throwing up the arms, namely, in response to the application of cold to the skin of the body During the first days the new-born baby is however strange this may seem—rather insensitive to all stimuli applied to the skin, be they heat or cold or pressure or the prick of a needle. But after the third week, if the baby is suddenly uncovered and exposed to a draft of cold
(136) air, or placed on a, cold linen cloth, or immersed in cool water, he may be seen to throw up his arms violently.
How, then, can these upward movements of the arms become associated, so to speak, with visual impressions? Incidentally the lands, during the upward movement, impress themselves as visual images upon the eyes,—upon the lower parts of the retinas, since all rays are crossed at the entrance to the eye. While the nervous process of the above mentioned reflex goes on, another nervous process thus starts from the lower region of the retina. The latter may be assumed to be especially intensive because it is caused by an object in motion. Although it is not perfectly known how this is brought about, it is a perfectly familiar fact that on the peripheral parts of the retina an object in motion produces a specially effective nervous process, much more effective than those resulting from the impressions of motionless objects. It therefore draws all the other visual nervous processes into its own channel and is the only visual nervous process which we have to take into account. The reflex motor effect of this process is probably a turning of the eye upwards. A considerable part of this visual nervous process is now likely to be deflected from its course by the other nervous process, still going on, into the direction of the motor organs which served the violent throwing up of the arms. Accordingly the resistance of the path leading over higher nerve centers from the lower part of the retina to the muscles raising the arm is reduced; and when this reduction of the resistance of the path has become sufficiently great, the hand will be stretched upwards in response to a visual stimulus coming from an object in front of the head and more or less above the eyes—a reaction which is quite common in the half-year old child. This variation of the nervous path is simply a
(137) further example of that kind of a "variation of response" in which the same response is called forth by a new kind of stimulus.
Of course, the establishment of an habitual upward movement of the hands in response to a visual stimulation does not necessarily interfere with the reflex of turning the eyes upwards in response to the same kind of stimulation. In order to establish the habit only a part of the nervous flux of the reflex need be deflected into the new motor outlet; and the final result may be a co-existence, a loose co-ordination of the two upward movements of the hands and the eyes. A similar loose co-ordination of eye and hand movements in a horizontal direction, to the right and left., can be observed in children as early as the third month.