The Fundamental Laws of Human Behavior
NINTH LECTURE
Max Meyer
Table of Contents | Next | Previous
Sensory condensation in piano playing, Proportional reduction of the resistance of higher and lower centers connecting the same corresponding points. The positive (and negative) susceptibility of a higher center greater than that of a lower center. Motor condensation in grasping.
We have studied and explained in detail one of the three kinds of variation of the nervous path, the simple variation of response. Let us study next that kind which we called sensory condensation. Musical practise may serve as an example. A child learning to play the piano, at a certain place in his piece, wants to strike two keys without having to look at both the corresponding notes in the score-no matter whether looking at both is possible or, because of their distance apart in the score, impossible. He begins by looking at one of the notes, seeing, let us say, the other one in indirect vision and therefore indistinctly. He reacts to the former by a perfectly definite finger stroke. But the other note, not clearly separated from its surroundings by the perceiving eye, calls forth an incorrect or insufficient movement. The child tries again, and now, of course, looks at the note whose movement was not properly executed. Everything is reversed. The finger movement which was previously done badly is now done well, and the movement which was previously done well
( 111) is done badly. After many trials, on the whole alternating as to the note favored by the eye, and as to the finger crowned by success, the child becomes able to perform both movements simultaneously with equal definiteness although lie is looking at only one of the notes-no matter which one lie is looking at. How do we explain this process of sensory condensation?
In order to explain it we make use of a diagram very similar to that of Figure 28, which helped us to explain the simple variation of a child learning to avoid contact with fire-. For simplicity's sake, we represent the stimulation by the note looked at as a single strong nervous process taking its origin from Sb (in Figure 34), and the stimulation
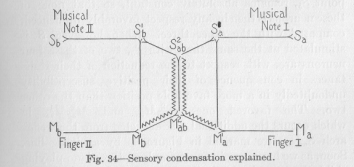
by the note seen indirectly as a single weak nervous process taking its origin from Sa. We learned from Figure 28 that the motor response to this double stimulation, strong at Sb and weak at Sa, occurs only at Mb. In the child's second trial everything is reversed. The stronger process is the one starting from Sa and the weaker the one starting from Sa so that the motor response occurs only at Ma. This goes on, the motor response occurring alternately at Mb and Ma. The question is what kind of changes the resistances of the various neurons undergo.
(112)
Plainly, when Sa and Sb are stimulated for the first time simultaneously and the motor response occurs at the point Mb (see Figure 28), the neurons S2ab M2ab and M2ab M1b suffer a slightly greater reduction of their resistances than in case Sb is stimulated alone. But it does not make any difference to the reduction of the resistance of the neuron S1b M1b , whether Sb is stimulated alone or Sa too. The flux from Sb divides at the point S1b and goes on to M1b partly over the direct, partly over the indirect route. Let us assume for the present, quite arbitrarily (we shall soon discuss this assumption separately), that the relative resistance of each of the four neurons S1b M1b , S1b S2b , S2ab M2ab , M2ab M1b in case the flux comes exclusively from the sensory point Sb, remains absolutely constant, so that none of these neurons is then in any respect favorably influenced, compared with the other three. When, however, Sa is stimulated at the same time with Sb, two of these four neurons are, with respect to the reduction of their resistances in consequence of their positive susceptibility undoubtedly in a more favorable position than the other two. The favored ones, S2ab M2ab and M2ab M1b , in which occurs the additional flux deflected from the reflex arch SaMa, are marked in Figure 34 by a zigzag line. Soon, as we have seen in our example of the piano-playing child, the reverse stimulation occurs. Sa is now stimulated strongly, Sb at the same time more weakly. Of the four neurons S1a M1a , S1a S2ab , S2ab M2ab , and M2ab M1a , two are now with respect to the reduction of their resistance specially favored, namely S2ab M2ab and M2ab M1a . They are marked in Figure 34 in the same way by a zigzag line. If these two events happen alternately (this is the important condition, without which the outcome would be a mere variation of response) in a sufficient number of repetitions, the final result can be read off
(113) from the diagram of Figure 34. The relative resistances will be gradually changed so that ultimately any flux starting from either Sa or Sb exclusively, will take, in the main, the path over S2ab M2ab, divide at the point M2ab almost equally, and cause a response at Ma as well as at Mb (a striking of both the piano keys), practically as if neither the neuron S1a M1a nor S1b M1b were any longer in existence.
Thus this kind of a variation of the nervous path sensory condensation-would have been explained, but only under the assumption just made that the relative resistance of any higher nerve center connecting the same corresponding points (for example Sb and Mb) as any lower nerve center remains constant, however much thé absolute resistance of all these centers may be reduced by a continuation or repetition of an isolated stimulation of the sensory point in question. This is the same assumption, only generalized, which we made with regard to four special neurons, and upon which we based our reasoning in the explanation of sensory condensation. This assumption is by no means self-evident. On the contrary, one might expect something very different. If Sb alone is stimulated, the flux in the neuron S1b M1b must be about three times as strong as in any of the neurons S1b S2b, S2ab M2ab, and M2ab M1b, since the path from S1b to M1b over these three neurons in series offers a greater resistance than the direct path over the single neuron S1b M1b. In consequence of the greater flux, the resistance of the neuron S1b M1b might be expected to be reduced, by and by, relatively much more than the total resistance of the other three neurons is reduced.
This last consequence, however, is biologically impossible. It would mean, in this special case, that the resistance of the neuron S1b M1b would tend to become so much red I aced. in comparison with any other nervous connection
( 114) between the points S1b and M1b , that the reflex arch would be practically separated from its connection with the points S2ab and M2ab. In general terms it would mean that all the reflex arches tend to become functionally separated from the higher nerve centers. Why should any animal have higher nerve centers, if the natural tendency of any nervous function from the beginning of individual life were to make the conduction of excitations over any higher centers more and more difficult and even impossible? The animal, then, might just as well, from the beginning of its life, possess no nervous conductors whatsoever, other than the reflex arches. It is clear, then, that we have, on quite general biological grounds, the right to assume that the relative resistance of any higher nerve centre connecting the same corresponding points as any lower nerve centre, in the case of isolated stimulation of the sensory point, remains constant. That is, in our special case (Figure 34), we have the right to assume, as we did, that when Sb alone has been stimulated, and the flux has occurred, in accordance with the length of the conductors, more strongly over S1b and M1b directly, more weakly over S2ab and M2ab, the ratio of the resistances of any two of the four connecting neurons involved always remains identical. Only simultaneous stimulation of a second sensory point can bring about a change of the relative resistances,-favoring the one or the other according to the special circumstances of the case. Our explanation of this case of sensory condensation is therefore complete.
The assumption made concerning higher and lower centers, which we found necessary on general biological grounds, may be stated also in different words. If the lower nerve center (take as an example the connecting neuron S1b M1b in Figure 34) does not, in spite of the
( 115) greater flux within it, suffer any greater reduction of its resistance than the higher center (S1b S2ab M2ab M1b), the positive susceptibility of the lower center is obviously less than the positive susceptibility of the higher center.
If this holds good for the positive susceptibility, we have reason to ask if it applies also to the negative suscep-tibility. That it does, seems to follow from the familiar factthat any ability is lost through the normal process of forgetting the less readily, the greater the number of instinctive, low-centered relations contained in it. For example, we forget how to recite historical dates more easily than we forget how to skate. Perhaps, then, we are justified in stating the assumption quite generally thus: The neurons of higher centers have a greater susceptibility than those of lower centers.
We have still to explain the third kind of variation of the nervous path, that kind which we called motor condensation. The condensation of the motor activity can be explained in practically the same way as a variation of response. We have learned from Figure 28 that, when Sa is rarely stimulated alone, but frequently together with Sb and then less strongly than Sb, the path leading from Sa to Mb tends to offer less and less resistance and finally to surpass greatly in conductivity the path from Sa to Ma. It is plain that when this stage of development has been reached, simultaneous stimulation of Sa and Sb can scarcely under any condition of relative intensity result in a simultaneous response at both I he motor points Ma and Mb, but exclusively in a response at Mb.
But not all cases of learning which deserve the name of motor condensation are so simple. Let us take as a concrete and not too complicated example the baby who, during several months, can grasp small articles only by closing
( 116) his whole hand, but afterwards learns to grasp a small article, for example a shirt button, between his thumb and index finger, like a grown person. At the sight of a sufficiently conspicuous object the young baby stretches out his arm. As a consequence of this reflex movement the fingers are stimulated by contact with the object. When the tip or any other part of the inner surface (from the nail toward the palm of the hand) of any finger is stimulated by touch, the finger reflexly bends. After a few months the baby learns to respond to the mere sight of an object by the initial part of the movementt of closing the hand; that is, the fingers begin to bend while the arm is still being stretched out, before they have had any contact with the object. What gives, at this stage, the grasping a particular appearance of clumsiness is the fact that the thumb does not aid in grasping, but is left practically functionless. To understand this is not difficult if we recall the statement just made that the original (reflex) bending of any finger results from touch stimulation of its inner surface. When the arm is stretched out and the hand naturally drops on the object presented, the thumb is not stimulated, as the other four
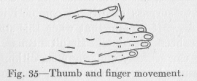
fingers are, on its inner surface, but on its side. This is the natural consequence of the anatomical location of the thumb, which moves, as the arrow in Figure 35 indicates, in the plane vertical to the plane of movement of the other fingers. Accordingly, the thumb has a less strong tendency to bend reflexly and, after the other fingers have bent, is prevented by them from bending.
( 117)
Compare with this clumsy manner of grasping the skill which the same child shows less than a year later, iii taking hold of a small object. While the arm is being stretched out, in response to the sight of the thing, the thumb and the index finger assume positions opposite each other, ready to take the object between them. The other three fingers remain all the time at rest, in whatever position they happened to be at the start. If the object is of medium size, the middle finger assists the index finger. Only if the object is large, do all the four fingers come into action. + In any case the thumb assumes a position opposite the fingers while the hand is still approaching the object, that is, in response to the mere sight of the object.
This whole process of learning can be made clear by the aid of a comparatively simple diagram. Sa represents, in Figure 36, the eye. The neurons S1a M1a and M2ab M1a leading toward the motor organ which stretches the index finger, are marked as not yet developed. The shortest motor outlet from Sa, is therefore Mb, the motor organ stretching the arm. In consequence of this stretching of the arm, the points Sd, Se, Sf, and Sg, representing the finger tips, are stimulated. The reflex response at Md, Me, Mf, and Mg is a bending of the four fingers and enclosing the object by them. The nervous processes from Sd, Se, Sf, and Sg attract, by dint of their own intensity, the process coming from Sa and force it to take largely the path over S2ab S3abcdefg M3abcdefg M2cdefg and thence into Md, Me, Mf, and Mg. The resistance of this path decreases until, after repeated occurrence of this deflection and the fixation of the path, the excitation caused at some time at Sa travels in the direction of Md; Me, Mf, and Mg and causes the beginning of a bending even before any of the finger tips have been touched.
( 118)
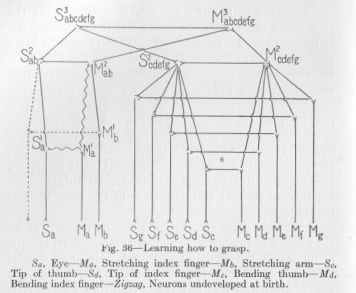
This accomplishment becomes the subject of a new influence at some time during the second half of the first year. The neurons marked in Figure 36 by zigzag lines reach by this time their full development and establish a new hereditary connection. When now the eye is stimulated, not only the arm but also the index finger is stretched out. This reflex appears at about the same time that the first articulated sounds (usually dental and guttural-"da" and "ga") are instinctively enounced by the baby,-a coincidence the significance of which we shall discuss at a later time. The result of this stretching of the index finger, with the other fingers in their ordinary slightly bent position, is touch stimulation of the tip of the index finger exclusively. Since the other fingers remain unstimulated, the index finger alone bends. If the
( 119) object is small, this bending movement of the index finger touching it is likely to pull the object toward the hand. When this is actually done, the object comes into contact with the outer surface of the other fingers and the inner surface of the thumb. The former contact is irrelevant, because it occurs on the outer surface; but the contact with the thumb is followed by a reflex bending of the thumb, so that the object is squeezed between the thumb and index finger. This means, in the diagram of Figure 36, that the two nervous processes from Sc to Mc and from Sd to Md deflect the nervous process coming from Sa by dint of their greater intensity and force it to take the path over S2abcdefg, M2abcdefg, and M2cdefg into Mc and Md. If this whole occurrence happens repeatedly, the resistance of the path from Sa into Mc and into Md is so much lowered that finally, at the appearance of the object, the excitation travels not only to Ma and to Mb, but largely also to Me and Md and causes a motor response here too, although slightly later than at Ma and Mb because of the difference of the distance. This makes it plain, why toward the end of the first year the child responds to the mere sight of an object by stretching out his arm and at the same time making the thumb and index finger ready to receive the thing which has been presented to the eye. The motor expansion, the clumsy use of the whole hand, has been succeeded by the motor condensation, the use of two fingers only.