Movement and Mental Imagery
Chapter 3: Movement and the Image or Centrally Excited Sensation
Margaret Floy Washburn
Next | Previous
IN the last chapter we found reason for thinking that a stimulus produces an effect on consciousness when it initiates a motor response which is partly checked in its execution by a process of inhibition, through the influence of an antagonistic motor response. The conscious process that is thus directly occasioned by the action of an outside stimulus is called a 'sensation,' and may be further distinguished as a 'peripherally excited sensation,' to denote the difference between it and a centrally excited sensation or image. For we can get, as conscious experiences, sensations not only from outside stimuli, but by the processes which are commonly known as 'memory' and 'imagination.' Not only can I see red when red light is acting on my eyes, but I can call up a mental image of red, and even, with fair accuracy, images of a whole series of different shades and tones of red. I can not only hear the tones of a violin playing the "Prize Song" from the Meistersinger when the violinist is actually before me (or the phonograph is actually running), but I can sit here in my study, with no actual sound stimuli acting on my ears save the voices of the children across the street, and hear the tones of the violin through the entire air. The very important question now confronts us as to whether these images or centrally excited sensations are each one of them dependent on an incipient motor process, as the corresponding sensations would be if outside stimuli were acting.
A fact clear to any observation is that there often intervenes, between the giving of a stimulus and the making of a movement that an outsider can see, a long interval. A man is sitting in his business office. To him there enters an acquaintance and
( 28) asks him to write a check for one hundred dollars. The man of business says nothing, and makes no visible movement for a considerable interval of time. His friend knows very well, however, that the request has been heard and is being pondered, and waits patiently. At the end of a certain period, the business man draws his check-book and his pen to him and carries out the request. He responds to the original stimulus by making the same movements which he might appropriately have made to it at once: but during the interval between stimulus and response. he will report from introspection, a train of processes has passed through his consciousness which had no outside stimulus: which belonged to the class of centrally rather than peripherally initiated conscious processes. He may have heard in memory the words of another friend urging the claims of the cause to which lie is asked to give; he may have had a mental picture of some scene from his past.
Now, it would be quite possible to hold (d) that while these conscious processes are, taken all together, the whole series of them, caused by the delay in responding to the original outside stimulus, and thus conditioned by the initiation of the final motor response, the several and individual centrally excited processes, images, or thoughts, that filled up the interval were not, each of them, dependent on an initiated motor response of its own. On the other hand, I think a very good case can be made out for the hypothesis (b) that each of these centrally excited processes, thoughts, or images, is dependent on its own special motor response. If the first view (a) is maintained, we should suppose that the energy of the stimulus S, not finding full discharge into the motor pathways of the response. passes directly through a series of sensory centres and finally, by this indirect route, finds its way back into the motor outlet which by the direct route was not fully open. As the nervous process traverses each of the series of sensory centres, there occurs, it would be held, a centrally excited conscious process in quality like the sensation which the centre in question would mediate if it were excited by an external stimulus. To take a simple ex-
(29) -ample: the words, 'I promised my wife to give this money,' may pass through the mind of the man who sits silently pondering between the request for the money and the writing of the check. According to hypothesis (a), the energy of the stimulus (the request for the money) passes directly through a series of auditory sensory centres, and the accompaniment in consciousness is the mental hearing of the words in question. The implication of this view is that every sensory centre may have its functioning accompanied by consciousness under two wholly unlike physiological conditions. The first condition is when the energy of an outside stimulus reaches the centre. As we have seen, it appears probable that in such a case consciousness results only when the motor response is partly but not fully produced; only when excitation is partly balanced by inhibition. The second condition is when energy travels to the sensory centre directly from another sensory centre. But if the mere passage of the nervous process from one sensory centre to another is sufficient to call up a conscious process; if, that is, the traversing of a sensory centre by a nervous process coming from another sensory centre and on its way to a third is sufficient to bring an 'image' into consciousness, why is not the passage of the nervous process through a sensory centre on its way to a free motor outlet sufficient to cause consciousness in the case of the peripherally excited process' Yet we have noted the probability that the traversing of a sensory pathway by the nervous process is unaccompanied by consciousness when the motor pathway is free. On hypothesis a, then, the conditions for consciousness produced by outside stimulation, on the one hand, and the conscious processes, 'centrally excited,' involved in memory and imagination, on the other hand, would be quite unlike: the former would demand not merely the traversing of a sensory centre by a nervous current, but the partial inhibition of a motor discharge: the latter would demand merely the passage of the nervous current through the sensory centre from another centre.
Other arguments against hypothesis a will present them-
(
30)
-selves later on. Hypothesis b, that each of the centrally excited processes which make up a train of images or thoughts has its own special motor response upon whose initiation it depends, may now be further developed.
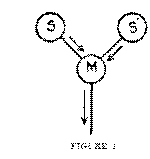
Suppose that a certain motor pathway, M, has at various times in the past been excited by energy reaching it from two different sensory pathways, S and S'. And suppose that in a given case energy from S reaches it, the effect of which is partly compensated by an inhibition from some antagonistic centre. M will then be in the kind of incomplete excitation which according to the conclusion of our last chapter is accompanied by consciousness of the sensation S. But whatever reason there may he for thinking that a process, accompanied by consciousness, is set up in S by the incipient excitation of M, would appear to hold also for the setting up of a process in S', a centre which is not now receiving any excitation from outside, but which has formerly, under the influence of outside excitation, discharged into M. The very same process which, on our hypothesis, when added to the effect of an outside stimulus makes that effect conscious, will, when it occurs in a sensory centre that is not being externally excited, be accompanied by the type of consciousness that we call 'centrally excited,' the consciousness that occurs in mental images and thoughts. When ever a motor pathway is at the same time excited by a sensory pathway and partially inhibited by an antagonistic motor excitation, a process occurs in all sensory pathways connected with the motor pathway by low synaptic resistances, including the sensory pathway that is exciting the motor pathway in question. This process is accompanied by consciousness. When it occurs in a sensory pathway that is being excited by an outside stimulus, it gives rise to the type of consciousness that we call a peripherally excited sensation. When it occurs in a sensory pathway that has no out-
(31) -side excitation, it gives rise to the type of consciousness that we call a centrally excited sensation or a mental image.[1]
How, under this hypothesis, can we explain the occurrence of a train of images or centrally excited processes, each one calling up the next? During the period between the getting of a stimulus to action and the carrying out of the action, a man's thoughts drift from one idea to another: he may in thirty seconds or so have a long train of conscious processes without any external stimulus. In the psychological laboratory, we ask students to take a certain word as a starting-point, and to introspect, without controlling, the images which follow one another through their minds.
On the theory which we are developing, such trains of centrally excited processes depend on the type of movement associations which we have called 'successive movement systems.' Take as an example the case of learning a series of nonsense syllables which are visually presented, printed or written, before one. For the sake of simplicity let us neglect the part played by auditory sensations, peripherally or centrally excited, and consider the process of pronouncing the syllables, as one looks at them, simply as a motor process whose stimulus is visual. In reading the series through we have a succession of visual stimuli, each followed by its proper motor response. Now, as the series is repeatedly read, each motor process comes to have two stimuli: the original visual one, the sight of the printed or written syllable, and also the kinesthetic 'feel' of pronouncing the syllable just preceding. The more thoroughly the series is learned, the more the kinesthetic stimulus becomes sufficient by itself, without the visual one, to set off the movement. When the series is thoroughly learned, one can say it 'without book,' that is, without any visual stimuli at all. If, however, in reciting a series of syllables without the copy before
(32) one, there is a delay; if some inhibition arises, then one may recall the look of the missing syllable, more or less accurately; that is, delay in the discharge of a motor centre produces, in the sensory centre which formerly discharged into it, that process upon which there is based consciousness of the sensation, centrally excited, which would be peripherally excited if an actual stimulus were acting on the sensory centre and its motor response were delayed.
This is what happens, we may suppose, when the series of syllables is actually being recited; that is, when the movements of articulation are being fully performed. If a hitch comes in the recitation, a mental image of the look of the syllables may be recalled. What, now, takes place when the series is not recited aloud, or even whispered, but merely 'run through' mentally% The thoughts or images which we have fancied to occur in the mind of the man deliberating whether he shall write a check do not take place, each of them, in the course of a series of actual movements that are visible externally. But nevertheless, — and this is a crucial assumption for our whole hypothesis, — there probably are, going on in his muscles, slight actual contractions. So when we mentally run over a series of nonsense syllables that we have learned by heart, it must be supposed that slight actual movements of the articulatory muscles do occur. Introspection furnishes some evidence of the fact. Try, for instance, the following test: pronounce aloud or in a whisper the letter ' b' successively twenty-five times, and as you pronounce it try, absolutely simultaneously, to think of each of the other letters of the alphabet. You will find that, whether you think of them in auditory or visual terms, whether you mentally bear them or mentally see them, you have to slip them in between your pronunciations of 'b': the sequence of events in your consciousness has to be 'h a,' 'b c,' 'b d,' and soon. Or try James's old experiment of holding the mouth open and thinking of the word 'bubble.' In this case, to my introspection, the auditory image is impossible, but I can form pretty well a visual image of the word. In general,
(33) visual images of words are not so closely dependent on articulatory movements, whether strong or weak, as auditory images are, for all visual images are related also to other movements, as for example movements of the eyes.
Slight but actual movements occur when we are thinking. These movements it is difficult to christen by a suitable name, but for convenience of reference, we shall call them 'tentative movements.' A later chapter will be devoted to their consideration.
A train of associated ideas, each one suggesting the next by a process which travels directly from one sensory centre to another, rests according to this hypothesis on a train of movements each of which, by the kinesthetic excitations its performance produces, sets the next one off, while delays in the process bring about, in sensory centres that have formerly discharged into the motor centres now partially excited, the processes on which the images are based. The case is fundamentally the same whether the movement system consists of movements that are being fully performed, though hesitatingly, or of tentative movements; whether the series of syllables is recited or only thought of.
Two points in favor of some such hypothesis may he noted. The older view demanded that there should exist nervous pathways directly connecting one sensory centre with another. Now, in those lower regions of the nervous system of vertebrates which have been the subject of exact physiological investigation and experiment, there is no evidence that sensory centres ever have pathways of direct connection with one another. 'Association pathways' connect an afferent or sensory pathway with an efferent or motor pathway. Several sensory pathways converge on a single motor pathway, their 'final common path,' or upon 'internuncial pathways' leading to a final common path, but nowhere does any path directly bridge across from one afferent path to another. Yet apparently such connections would be as useful in the lower regions of the nervous system as they are supposed to be in the upper, cortical
(34) regions. Simplicity would argue for the supposition that all parts of the nervous system are built on —the same general plan, and that this plan involves the association of movements, but not the direct association of sensory processes.
A second argument for the kind of hypothesis we are trying to form is this: The older view supposed not only that direct pathways connected one sensory centre with another, but that when the two sensory centres were simultaneously excited, the resistances along the connecting pathway were lowered, so that at a later occasion, when a nervous process occurred in one of the centres, it travelled across to the other centre and 'centrally excited' it. Thus was explained the fact of experience that in order for one thing to recall another to our minds, by central excitation, we must have encountered the two things together. In order that the sight of a word printed in a foreign language shall recall its correct pronunciation, we must have heard the word pronounced on some former occasion while we were looking at it. But the older view neglected the fact that merely experiencing two stimuli together will not suffice to form a central excitation tendency. Stimuli may easily act simultaneously, each on its own sensory pathway, and the resistances between these pathways not be lowered at all. For example, one needs merely to suppose that when looking at the foreign word for the first time and hearing it pronounced, one hears it only inattentively. If the attention is not directed towards the sound, if both the look of the word and the sound of it are not attentively experienced, the look will not later recall the sound, nor vice versa. Not the simultaneous experiencing of two objects, but simultaneous attention to them, forms a tendency to recall.
It is clear on our theory why the formation of an 'association' demands attention. All association is association between movements. Now, when the association between movements is in process of establishment, the performance of the movements is necessarily subject to many inhibitions and delays, but of course it must be an actual performance. We must
(35) suppose then that the actual performance of movements — that is, something more than the mere innervation of the muscles —is what is involved in attention; the excitation being weakened and slowed by inhibitions, but nevertheless resulting in actual though slight movements. We have seen that there are two types of association between movements. The first type is characterized by the dropping out of movements, so that a stimulus takes on a new motor response and loses its old one. The second type preserves all the movements, and links them so that the performance of one furnishes the stimulus for the performance of another. Clearly, it is this second type of association that is the basis of trains of ideas, each one distinguished from the rest, for only where different motor responses are initiated are the sensory processes leading to them accompanied by distinguishable sensations.
One of many other points in favor of a motor theory of attention and the image is that it explains readily what we may call the transitory character of both. "The object of attention constantly fluctuates." We cannot hold an unchanging thing in the focus of attention: it seems to change of its own accord. Some years ago in the Vassar laboratory (124) an attempt was made to investigate experimentally a phenomenon that nearly every one has noticed: the fact that if you look steadily for some time at a printed or written word it takes on a strange and unfamiliar appearance. As our observers gazed steadily at a single printed word for three minutes, they noted that the word was constantly changing; constantly suggesting new pronunciations, new syllable divisions, and so on. 'Acre' would become 'ac-er,' 'a-cree,' and so on. This shifting of the object of attention has been referred to the fact that primitively a stimulus must be a change in the environment of an organism, and that an unchanging object cannot be a stimulus. But in fluctuations of attention it is not really the stimulus, not the external object, that changes. What changes is our reaction to it, and the reason for this constant change of response to a stimulus in attention is that the essence of attention is movement, and
(36) that the great majority of motor processes are of the phasic rather than the static or attitude type.
On our general theory, then, consciousness will depend on a certain ratio between excitation and inhibition of the motor response to a stimulus; while recall through association will depend on the motor response's being sufficiently innervated to be actually performed, though only in the weakened form of a tentative movement. We may tabulate the different ratios of inhibition and innervation of response which according to this theory correspond to different degrees of consciousness.
I. Motor response very slightly excited, not enough to produce consciousness. The effects of this excitation appear, however, in the phenomenon of 'readiness,' which will be discussed later. (gee page 82.)
II. Motor response more strongly initiated. but no actual performance of a movement, either tentative or full. This is accompanied by consciousness of the stimulus, but no associative activity can take place, because associative activity requires the actual performance of a motor response, to produce the kinaesthetic excitations necessary to set other motor pathways into action.
III. Motor response still more strongly initiated. Tentative movements occur slowly and with delays. Consciousness and associative activity both present: this is attention.
IV. Motor response still more strongly initiated. Tentative movements occur smoothly and without delay. Associative activity occurs, with comparatively little 'conscious accompaniment: these are the conditions which we may think of as underlying very rapid thinking, which is nearly unconscious.
V. Motor response fully initiated and entirely unopposed. Performance of full motor response without delay and without consciousness: the situation in secondarily automatic or habitual actions.
Thus, by supposing five different ratios of excitation to inhibition, we can make a bridge between the apparently con-
( 37) -tradictory but equally true statements that unconsciousness accompanies total inhibition of movement and that it accompanies perfect freedom of movement.
Can we form any notion of what physiological process underlies the difference which we consciously experience between a peripherally excited sensation and an image? We have assumed that the degree of consciousness, what M�nsterberg would call the vividness, of both is due to the same underlying cause, the relation between the excitation and the inhibition of the motor pathways with which their sensory pathways are connected. Now, just what makes the difference between the color red as I see it before me and the color red as I remember or imagine it? Certainly it is not a difference in consciousness degree. The degree of consciousness depends on attention: but I may be wholly inattentive to the colors I am actually looking at, and fully absorbed in the color imagery that my imagination conjures up. If you ask me afterward what I thought of a certain red cushion, I may have no recollection that any such object was near me, although I spent some minutes apparently looking intently at it.
All authorities are agreed that centrally excited sensations are much less steady and enduring than peripherally excited sensations. I cannot hold in consciousness the image of red nearly so long as I can be conscious of the red that I actually see. On our theory, this short duration of the image is due to the fact that the stimulus which initiates the motor response on which consciousness of the image depends is produced by another movement; while the stimulus which initiates the motor response on which the peripherally excited sensation depends is an outside physical force. It is obvious that a stimulus supplied by a bodily movement must be of brief duration, unless indeed the movement is not a true movement, but a static motor process or attitude. Much discussion has centred about the problem as to whether a difference in intensity is characteristic of the difference between a sensation and an image. To immediate observation it seems clear that the color or tone or
( 38) smell one imagines is never so intense as an actual sensation. Now, it is not so easy as one might suppose to decide exactly what is meant by intensity.
Schaub's (122) observers, in an introspective study of the intensity of images, made the following reports: the images "lack volume, that is, concomitant muscular and organic sensations": "with the sensations there are kinaesthetic accompaniments which are not present in the image"; "images exactly like the sensations in intensity, but ... they did not give the kinesthetic shock that accompanied the stimuli." There seems introspective evidence that when an outside stimulus acts on the organism, it sets up, not only a definite motor response that belongs peculiarly to it, but also a more generalized and diffused motor response, an 'all over' disturbance. This general disturbance is felt in consciousness sometimes as fused with the sensation proper to the stimulus, if the kinesthetic and organic sensory excitations which it sets up do not produce a special motor reaction of their own; sometimes it is attended to on its own account, and more or less analyzed into localized components. Whether analyzed or unanalyzed, the kinesthetic excitations coming from this general motor disturbance are probably the basis of a characteristic difference between the sensation from an outside stimulus and the recalled image; of what Ziehen (161) has termed the "sensual vivacity" of a peripherally excited sensation, and Stout (134) calls the "aggressive character" of a sensation as compared with an image. It is true that Stout, although he admits that when an outside stimulus acts, "the whole organism receives a shock giving rise to a mass of organic and kinaesthetic sensations," expressly denies that these can be responsible for the aggressiveness of the sensation. But his objection does not seem well taken. He says: " It seems evident that they [the kinaesthetic and organic sensations] cannot give an aggressive character to the experience unless they possess this character themselves, and as a matter of fact they are highly intrusive and obtrusive. But if organic sensations can 'strike the mind'
(39) in this way, there is no reason why other sensations should not do so too. The ultimate appeal must be to introspection. This shows in the case of the steam whistle [the example Stout has been using] that the sound itself is aggressive in the same way as the organic sensations which accompany it." But this reasoning confuses an unanalyzed experience with an analyzed one. We may reply that the so-called 'aggressiveness' of a sensation is merely the name we give to the unanalyzed 'feel' of the general organic shock produced by the stimulus; when this 'feel' is analyzed and certain sensational components are attended to by themselves, they in turn may perfectly well be accompanied by a diffused motor response in addition to the special motor response involved in attending to them; if they are so accompanied they also are felt as 'aggressive.' The same answer may be made to Stout's further suggestion that, "The organic sensations follow the beginning of the sound after the lapse of about a second, but the sound itself is aggressive from the outset." It is the analyzed experience that is thus delayed; not the unanalyzed feeling of aggressiveness.
When an actual outside stimulus, then, is responsible for the production of a sensation, the sensation has a certain vivacity or aggressive character that I shall assume to be due to kinaesthetic and organic excitations resulting from a general motor discharge. This character is quite evidently stronger, and the general motor discharge is more marked, the greater the amount of the stimulus: there is no reason why it should not be identified with the character of intensity. Shall we, then, say that images have no intensity?
Such a general motor response may be produced, like any other movement, not merely by an outside stimulus, but also by the setting up of another movement: that is, it can enter into a movement system. Thus, I can imagine a very loud sound, and when I do so, I feel certain sensations of shrinking and shock which are evidently the result of movements produced because they are associated with the motor processes involved in the idea, 'very loud sound.' They are associatively
(40) produced, and not caused by the action of an outside stimulus on my ear. An image may be more or less intense (as distinguished from being more or less clear, having more or less consciousness degree) according to whether these sensations correspond to a more or less diffused shrinking and shock. In this sense images may possess intensity: I may have the image of a louder or a weaker sound. But introspection seems to testify that the general motor responses thus associatively produced are not identical with the general motor responses that the original outside stimulus produced. The general shock and strain felt when I imagine a very loud sound are like but not identical with those I feel when I actually hear the sound. The difference very probably consists in the fact that the associatively produced shock involves merely the kinesthetic and organic sensations which were analyzed out of the original, and that what is missing is the unanalyzed and more diffuse components: it would naturally be the components which were attended to separately that could readily enter into a movement system and so be associatively produced. In any case, while the intensity of a mental image consists of the kinaesthetic-organic 'feel' of a general motor response, the 'feels' of which image intensities consist are not identical with those of which sensation intensities consist. Thus, it is possible for Ziehen to say of "sensual vivacity" as a feature of the peripherally excited sensation that it " does not belong at all to the idea, not even in a diminished intensity," and for Schaub's observer to state that the image was just like the idea in intensity, but did not give the same kinesthetic shock.
Before we proceed to a fuller discussion, much needed, of the subject of tentative movements, it will be well at this point to consider a little further the state of affairs described under II, above: the case where the motor response is very slightly initiated, not enough to produce any actual muscular contraction, either tentative or full. This, it was said, produces consciousness of the stimulus, but under such circumstances the sensation resulting can suggest nothing associatively. Now,
( 41) suppose that a number of stimuli are simultaneously acting on the organism, and that each of them is exciting its appropriate motor response to precisely this degree. The result for consciousness will be a fusion or blending of sensations which is recognizably a complex phenomenon, but is not analyzed. We may say that when the conscious state at a given moment is felt to be complex and not simple, even though the complexity is not analyzed into its component parts, the reason is that a number of different motor responses are very slightly initiated. As soon as one begins to analyze such an experience, one of course attends to its parts successively, and the motor excitation belonging to each part attended to passes from stage II to stage III; to the stage, that is, where it becomes an actual tentative movement. Thus, for example, we hear a musical chord. It makes upon us a complex impression, even before we analyze it into its constituent tones; this complexity is due to the fact that a number of different motor responses are very slightly excited. When one analyzes the chord, one gives attention to its tones successively, making tentatively the motor response belonging to each.
All unanalyzed complexity of experience depends on the simultaneous initiation of different motor responses, none of them carried to the point of actual performance, even as tentative movements. A fine example of the truth of this statement is furnished by the phenomena of single and double images in binocular vision. When you hold up one forefinger about a foot from the eyes, and look at it, the other forefinger, if held at either a greater or a less distance from you, will appear double. Now, according to our principle, a single object is one which initiates a single movement; an object can be perceived as two only if it initiates two movements. The law under which objects are seen singly by the two eyes is generally stated as follows: if the images of the objects fall on corresponding points of the two retinas, they will blend into one; otherwise they will produce double vision. Corresponding points are points that would coincide if one retina could be placed directly over the
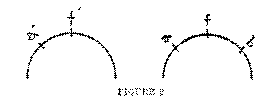
(42) other; thus, evidently the centres of the two retinas are corresponding points, and so are any two points that lie at equal distances from and in the same direction from the centres of the two retinas. -Now, it is clear that when the images of an object fall on corresponding points, they suggest absolutely identical movements of the two eyes. The movement that regularly tends to be initiated when an image falls on a point on the retina is the movement which would be required to bring the image from that point to the centre of the retina, since it is when an object casts its image on the centre of the retina that it is most clearly seen. If, for example, the images fall on the corresponding points a and a', the movements a — f and a'—f' are initiated, movements precisely equal in extent and direction: practically a single movement of the two eyes. But if the images fall on points a and b', non-corresponding points, then the movement a — f will be initiated for one eye, and the movement b' — f for the other eye. These are movements in opposite directions, unlike movements; hence, may we not say, the object is seen as double?
Two further considerations may be noted. First, on our theory of the nature of the associative process, evidently two sensory centres or pathways must be connected with one and the same motor pathway, to give rise to a centrally excited sensation or image. For the centrally excited process is due to the partial excitation of the motor pathway belonging to the sensory centre which is centrally excited, through the action upon it of another, a kinaesthetic, sensory pathway. In the diagram, on page 30, we may assume that the sensory centre S is kinaesthetic. The excitation of M, meeting a certain amount of inhibition, sets up a process in S', which at some former time has discharged into M; the accompaniment of this
( 43) process is a centrally excited sensation or image of the proper stimulus of S. At the same time, according to our theory, there should be peripherally excited consciousness corresponding to the kinaesthetic pathway's excitation. Thus, in every centrally excited or image process there would be two components, one kinaesthetic and one of the modality to which the image is referred; in every centrally excited visual sensation, for instance, there would be a kinaesthetic and a visual factor; in every centrally excited auditory sensation a kinaesthetic and an auditory factor. These factors need not, however, be for consciousness in any way distinct, since distinctness for consciousness involves the excitation of two different motor responses, and the kinaesthetic and non-kinaesthetic components of the process underlying the image need not give rise to different motor responses. The interesting point, however, is this. As the associative processes run more and more smoothly; as thinking becomes easier and easier along a familiar line, as the motor responses pass from stage III to stage IV, and the delay becomes less, it is easy to suppose that the non-kinesthetic pathways become less and less involved, and that the conscious accompaniment of thinking becomes less and less composed of centrally excited visual, auditory, and other non-kinaesthetic processes, and more and more composed of peripherally excited kinesthetic processes. Thus we may be able, in a later chapter, to understand the fact that thinking tends to become 'imageless' in proportion as it becomes more rapid.
Secondly, the most natural explanation, on a theory which bases the image on the movements made in connection with attention, for individual differences in type of imagery is that they depend on individual differences in the appeal of certain kinds of stimuli to attention. A person thinks in visual imagery habitually if he habitually attends to the look of things; he thinks in auditory imagery if he habitually attends to the way things sound; he is clearly aware of his own tentative movements if he is in the habit of attending to movements. What determines the habit of attending to visual stimuli more than
( 44) to auditory stimuli may be either the nature of one's occupation, or the development of one's sensory apparatus. A person whose eyesight is not good would not be expected to have his attention readily attracted to the appearance of things, and would probably not belong to the 'visual type' as regards his imagery; if, however, with the aid of eye-glasses he were a great reader, his visual imagery for words would probably be far in advance of the rest of his mental pictures. A person with keen eyesight. whose profession did not involve the use of visual imagery, would probably have much less of it than, say, an architect or a painter. A person who has poor discrimination for musical tones and intervals, that is, whose auditory sensory apparatus is poorly developed, would have less auditory imagery for that reason, although he might increase his powers of imaging sounds if his profession demanded attention to sounds. 'Imagery types' and their variations and anomalies may, I think, be most readily explained as attention types, due to innate differences in sense discrimination and to habits formed by the nature of one's environment and work.
It is evident that this theory of the motor basis of consciousness, whether centrally or peripherally excited, requires, for a creature like man, with an immense variety of sensory discriminations, a great variety of movements. Every stimulus, in so far as it is discriminated from other stimuli, excites a motor response different from that which other stimuli excite. It is possible, indeed, that two stimuli may be consciously discriminated and still produce the same externally visible movement: a person may reach for two different kinds of food with equal vigor and enthusiasm and yet be perfectly aware of their difference. But the motor reactions in such a case are complex, and, while outwardly alike, the inner components may well be very different: the person, for instance, may be mentally naming the two kinds of food. If the whole motor response were the same for two kinds of stimuli, then our theory assumes that they would not be in any sense discriminated. To have a variety of sensations and images, then, demands a variety of
( 45) movements. Let us very briefly survey our motor stock in trade.
The muscles of the limbs acquire very few new movement systems in the course of a lifetime, and their movements have little variety. Those of the hands, however, are quite another matter. The most complicated and varied kinds of finger movement systems that we acquire are of two types. There is, first, the type where an object is held between the thumb and one or more other fingers, and moved in various directions. These are the movements which we use in forming written letters, holding a pen or pencil; in drawing, in using delicate tools. They form successive movement systems; in the case of writing or drawing, the movements leave behind them permanent visual stimuli, so that visual imagery tends to be lastingly associated with them. Secondly, there is the type of movement which consists in pressing down with the finger tips, using different fingers successively. In typewriting common words like 'and,' 'the,' and many others, firmly established successive movement systems are formed, and in learning a piece of music by heart the finger movements involved in various phrases become well organized into systems.
The muscles of the face have a certain number of innate movement systems. These were originally all phasic systems, it may be supposed, giving rise to changes of facial expression in connection with various stimuli of importance for welfare: Wundt has shown how, by the operation of the principle of analogy, movements originally connected with accepting and rejecting certain smells and tastes have become associated with other agreeable and disagreeable stimuli. It is noteworthy that facial expressions tend to become static rather than phasic. An expression originally appropriate to a temporary emotion becomes fixed and permanent. The reason for this is probably that the facial muscles are little needed and may remain at rest for long periods of time: a phasic system of arm movements, obviously, could not afford to become static, since the muscles would be called into activity in other combinations.
( 46)
The eye muscles are an extremely interesting system from the point of view of the movement systems into which they enter. The fixation of the eye in any position may involve a simultaneous static movement system; that is, a true attitude, requiring the steady contraction of certain muscles. The unopposed contraction of the external rectus muscle, for instance, would move the eye away from the nose until the limit of the muscle's contraction was reached: if any point between the centre of the field of vision and the extreme side is fixated, the contraction of the external rectus is balanced by that of the internal rectos. and the contractions have to be maintained as long as fixation is maintained. Binocular fixation also means a static movement system, involving varying degrees of steady contraction on the part of the two internal recti, acting together and producing convergence of the eyes. The eye movements by which lines are explored are, as we have seen, while not successive movement systems themselves, the basis of the formation, between the other movements made in response to visual stimuli, of double successive systems with reversed order, the characteristic basis of spatial perceptions.
The articulatory muscles furnish us with the greatest variety of movement systems, a variety that is practically unlimited. We have only to consider that it includes at least all the words and phrases that can be uttered in any language spoken on the earth's surface, to realize its enormous range. There is nowhere else in the body a region capable of anything like such versatility: the movements of the fingers, complicated as their combinations may be, have not the same varied character to start with; their combinations must be formed out of a much smaller number of simple movements. In speech the single sounds are produced by means of simultaneous phasic systems. The sound of broad a, for example, results from the simultaneous vibration of the vocal cords and a certain position of the cheek muscles and the tongue. The guttural g is produced by a certain position of the glottis and a slight vibration of the vocal cords. A word involves a successive system made up of the
( 47) simultaneous phasic systems that constitute the letters; a phrase, a sentence, a stanza of poetry learned by heart, are still longer successive systems. The slight degree of physical effort which the use of speech muscles involves, and the fact that they are not often required for ministering to immediate practical needs, makes possible the repetition of a given movement system so often that it may be very quickly and firmly established. Another characteristic feature of articulatory movements is that they so commonly produce audible effects, which for a given movement are absolutely constant in character. Very commonly as a successive system is formed out of speech movements, the auditory pathways excited by the performance of the first movement join with the kinaesthetic pathways in exciting the second. This is surely the reason why kinaesthetic excitations from speech movements, if accompanied by consciousness at all, seem to involve some auditory imagery.
Of the greatest significance for the structure of our experience are undoubtedly the motor processes which go on in the internal organs. The general organic response to a stimulus furnishes masses of kinaesthetic excitation which we cannot analyze in many cases, but which are probably highly varied and capable of representing. in movement systems, the different individualities of many stimuli and combinations of stimuli. The possibility that the peculiar motor effect of a stimulus on the organism as a whole may be the basis of the recall of mental imagery has not escaped psychologists. Betz (11), in particular, says that one experience recalls another when they set up the same Einstellung or attitude, and this Einstellung seems to include the total motor effect of the experience, not only word movements, but "finer organic movements," and feelings. As an illustration of an Einstellung which in a given case accompanied the recognition of a face he mentions the tendency to laugh. M�ller-Freienfels (9e), also, makes the 'attitude' into which an object throws us, and which involves apparently as its most characteristic features organic processes, the essential thing for both perception and recall.