The Fundamental Laws of Human Behavior
TWELFTH LECTURE
Max Meyer
Table of Contents | Next | Previous
Repetition of motor activity characteristic of learning in childhood. Variation of the order of earliest accomplishments. Speech organs: whispering and singing. First speech sounds. General and specific resistances of neurons. The motor outlet of a group of successive nervous processes determined by the temporal order of the qualitatively different processes. The general resistance as well as the specific resistance reduced by any special flux; but the reduction of the specific resistance outlasting that of the general resistance. Possibility of a particular distribution of specific resistances resulting from experience.
OF the motor activities of early childhood we have briefly discussed thus far the adaptive movements of the sense organs of the head and the fundamental activities of the hands and feet. All the processes of learning which we mentioned can easily be placed in the classes distinguished by our theory of nervous activity. Most of them are simply variations of response, some are kinds of condensations. That the learning process is so quick in all these cases is largely due to the fact that the reflexes in question quite naturally lead either to an exact repetition of the stimulus or to a stimulus similar to the first and even more powerful —as when a baby piles up blocks. Even when a new stimulus caused by the first reaction is dissimilar to the
( 154) first, it may take the path to the same motor point, either according to the law of deflection, or because of the temporarily lessened resistance in that direction, or owing to both these causes. It has become customary to refer to these repetitions of motor activity by the term "circular reaction.' Such a special term is useful as it emphasizes the typical character of learning in early childhood in comparison with learning in later life, when the conditions are more complex and repetition of the motor activity rarely results. But let us not forget that the term "circular reaction" is not itself an explanation and would be an additional mystery unless we explain the facts themselves as we have very briefly attempted to do.
Let us emphasize that, whenever we spoke of a particular kind of movement as preceding in human growth another kind, we wished to indicate merely the most common serial order of development, and that individual deviations from this order, displacing relatively for four or five months this or that element of action, are common too. Parents should neither be alarmed nor think that they have special reasons for paternal pride if they observe that their children do this or that several months later or earlier than other people's children, or that similar differences appear among their own children. Human life during the first—and even second—year is so elementary, that any conclusions with respect to the enormously complex life of the future have as much value as the oracles of antiquity.
The new-born baby is so helpless a creature that, if experience did not teach the contrary, one could hardly believe that such a being could survive the accidents with which he must be daily confronted. When he has reached the age of fifteen months, it seems much less improbable that he will succeed in the struggle with the world. He
( 155) has learned to use his sense organs, he has acquired the ability of locomotion which lie daily improves by adding further skill in walking, running, and climbing, and he has learned to use his hands so that only further exercise is wanted to enable him to shape the world in accordance with his needs. Yet, at this age it is still easy to recognize that all the diversified human power is merely a complcx of a small number of natural laws because we can still trace the child's power back to its sources. Of course, the few muscular activities which we have mentioned as characteristic of the behavior of the infant, are not really so simple as we have—intentionally—represented them, but are movements executed each by a large group of muscles and permit each many modifications, according as the one or the other muscle in the group receives the stronger nervous impulse. During the second year the complications grow almost into the infinite, especially during the second half of this year, when the child learns to use more and more effectively its speech organs. Nevertheless, we shall convince ourselves that even now nothing happens but what can be derived from the laws of nervous activity previously stated and a few additional laws which we shall state below.
It may be well here to make a brief statement as to what the essential factors of speech production are. There are two main classes of sound producing organs. The first class contains only the pair of vocal chords in the throat, at the end of the wind pipe. The vocal chords are comparable to an ordinary musical wind instrument. They produce, when the mouth organs leave the out-streaming air unobstructed, pure musical tones. With-out the vocal chords one cannot sing. With the vocal chords in a normal condition, one could sing even though the mouth organs were completely lacking. The vocal
( 156) chords might therefore be called the singing organ. On the other hand, the totality of the month organs, including even the nasal cavities, might be called the whispering organ. We can make ourselves understood even though we may be unable to sing; for example, when we are hoarse, we can still whisper. Our ordinary speech is a compound of whispering and singing, only that the latter, the "song" of speech, is not, in a musical sense, melodious.
The first cry of the baby, immediately after birth, is a song, in the sense n which we have just used this word. It is only two or three months later that the growth of the nervous system has progressed far enough to make possiblethe reflex production of speech sounds. Naturally, first those speech sounds are called forth, which require the smallest amount of sensori-motor activity, of muscular contraction. These are the guttural sounds, produced by the mouth organs located farthest back, near the throat, and the dental sounds, in the production of which the teeth (or gums) co-operate with the tongue. Neither guttural nor dental sounds require the lips. It is natural again, that of the various guttural and dental sounds those are produced first which require the smallest amount of muscular work. We are therefore most likely to hear first such sounds as ga or goo and da or doo, later the sounds of k, t, etc. The accompanying vowel depends, in accordance with the laws of acoustics, on the measurements of the cavity which the mouth happens to form at the moment. Labial sounds occur several months later, since the tension of the lips requires more muscular work: we then begin to hear b and m and nasal n. The popular notion that the first words pronounced by a baby are papa and mama, is an illusion.
One of our tasks in connection with speech is to explain how stimulation of the ear by different words, for example,
( 157) by the syllables ga and da, can call forth definite and different motor responses not only of the speech organs, but of any motor organs of the body. Here we are con-fronted by a difficulty which hitherto, in order to avoid complication of the discussion, we have intentionally evaded, but which we must now face. The question, Why does excitation occur at a definite motor point? we have always answered thus: Because over the path from the definite sensory point excited to this motor point the nervous flux finds less resistance than on any other path. Now, it. is plain that stimulation of the ear by the syllables ga or da can not mean the stimulation of two different sensory points. It is true that the theories concerning the physiological processes in the ear show wide differences of opinion; but that the difference of da and ga is simply a local difference of sensory points from which the two nervous processes start, no physiological theory is likely to assert. That a nervous flux takes the path toward one motor point rather than toward another, must depend, not exclusively on the sensory point whence it comes, but sometimes also on qualitative differences of the flux itself. The importance of the role played by qualitative differences of nervous processes becomes still more apparent when we consider the fact that words differ, not only in the quality of the various stimuli, but especially in the temporal order in which the sounds follow each other. For example, the main difference between the words cat and tack consists iii the difference of the temporal order of a small number of sound qualities. There is no doubt that such non-spatial differences of stimulation are sufficient to bring; about spatial differencesin the motor response.
In order to explain these facts, it is not enough to speak of the resistance of a neuron. We must speak of a general
( 158) resistance and specific resistances to particular kinds of flux. How is it possible for a single conductor to have several specific resistances? We can understand this if we assume, as we have previously done, that a nervous flux is a wandering of ions. In a highly complex chemical substance like that which makes up nervous tissue, many kinds (hundreds or even thousands) of molecules may serve as ions. Our purely mechanical analogies, of course, become insufficient at this point, for, as soon as we assume different kinds of ions, we would have to use as analogy a stream of which not always the whole substance, but sometimes only these, sometimes only those of the particles composing the substance may be in motion, while other particles uniformly distributed through the whole substance may remain at rest. This is something like the streaming in filtering and in osmosis, but even more complicated than such processes.
The assumption of specific resistances of any neuron is a brief expression of the fact that the motor outlet of a nervous process may be determined by the quality of the flux. Now we must find a way of expressing by a picture the fact that the motor outlet of a definite group of processes quickly succeeding each other may be determined by the particular temporal order of the qualitatively different processes, in order to make plain that, for example, the response to the word cat may and does differ from the response to the word tack. Let us use for this purpose the diagram of figure 42. In this figure S0 represents a peripheral sensory point; S1 a central sensory point from which several neurons pass on in different central directions; M2abcand M2def central motor points from each of which numerous neurons pass on in different peripheral directions. Imagine that the words cat and tack consist of two sounds, let us say, ca and ta, and that the difference
( 159) of the two words lies in the order of succession of these two sounds. While this is only very approximately true, the simplification thus brought about is justified by our purpose. Suppose that the nervous process belonging to the sound ca is x, that the nervous process belonging to the sound ta is y, and that in Figure 42 the neurons S1S2abc, S2abc M2abc, M2def M1f and M1f Mf have an extra-ordinarily low specific resistance for the flux x (as indicated
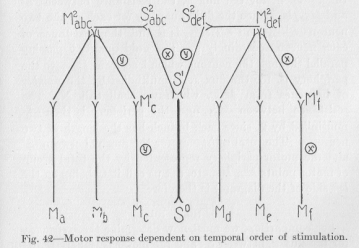
in the figure by the letter x inclosed in a circle) and the neurons S1S2def, S2def M2def, M2abcM1c, and M1cMc , an extraordinarily low resistance for the flux y (as indicated by the letter y inclosed in a circle). Our task is to explain why the motor response occurs at Mc when from Sº the flux x precedes the flux y, and at the different point Mf when the temporal order of x and y is reversed, to show that this difference of response is possible. We shall show it in exact mathematical terms.
(160) Suppose the fluid quantity passing through one neuron in a unit of time to he equal to G in case general resistance is effective, equal to S in case specific resistance is effective; and that in the latter case the flux is greater, so that S is larger than G, perhaps a multiple of G. Suppose further that the resistance between two "levels" of the nervous system distant the length of one neuron is equal to the reciprocal value of the fluid quantity passing in a unit of time. Then, for the flux x, the resistance between S1 and M2abc is 2/S since the fluid quantity for each of the two neurons S1S2abc and S2abcM2abc is S and the resistance of each neuron accordingly 1/S. The fluid quantity passing parallel through the neurons which lead from M2abc to the periphery is, for the flux x, G multiplied by the number of branches from M2abc, which we shall call N. The resistance between M2abc and the periphery is then 1/NG multiplied by 2, since each branch has the length of two neurons. The total resistance, for the flux x, between S1 and the motor periphery represented by the total group of motor points Ma, Mb etc. (let us call it L, being on the left side of the diagram) is therefore
L = (2/S) + (2/NG)= (2(NG+2))/NGS
For comparison, we must now determine the resistance, for the flux x, between S1 and the motor periphery represented by the total group of motor points Md, Me, etc. (let us call it R, being on the right side of the diagram). The resistance between S1 and M2def is 2/G. The resistance between M2def and the motor periphery is to be found as follows. The fluid quantity passing parallel through the neurons which lead from M2def to the level one neuron nearer the periphery, is S plus (N-1)G, since one of the neurons branching off from M2def has for x a specific resistance, 1/S, and the (N -1) other neurons have the general
( 161) resistance 1/G. The resistance between M2def and the periphery is then the reciprocal value 1/(S+(N-1)G), to be multiplied by 2 since each branch has the length of two neurons from M2def to the periphery. Consequently:
R=(2/G)+(2/(S+(N-1)G) = 2(S+NG-G+G)/G(S+NG(-G) = 2(S+NG)/G(S+NG -G)
In order to compare the magnitude of L with that of II, let us form the ratio:
L/R = 2(NG+S) X G(S+NG-G) / NGS X 2(S+NG) = (S + NG - G)/ NS
Let its consider this formula in a few special eases. It appears that, when there is only a single motor branch at M2abc as well as at M2def, that is, when N equals 1, L attains its maximum value, is equal to R, for
L/R = (S+G-G)/S =1
With increasing N, the ratio L/R grows smaller, for each unit added to N is multiplied in the numerator only by G, but in the denominator by the larger value S. If N becomes a very large number, the ratio
L/R = (S + NG - G)/ NSapproaches more and more G/S, which is the lower limit of this resistance ratio, for in the numerator the constant value (S-G) becomes then infinitesimal relative to NG, so that the ratio reduces to
limit (L/R) = NG/NS = G/S < 1We see, thus, that the advantage of the left side of the conductive system of Figure 42, which is equivalent to smallness of the resistance ratio L/R , for the flux x, depends on two conditions, (1) on the degree by which the specific resistances is reduced below the general resistance 1/G, or, in other words, by which the flux intensity S surpasses
( 162) the flux intensity G, (2) on the number N of the specific motor points belonging to the system. Now immediately after the termination of the flux x, the flux ;y takes its origin from the same point Sº. In order to give x, in this theoretical discussion, no unfair advantage over y, we have supposed our conductive system of Figure 47 to offer to y all those resistances in the direction of "left-right " which it offers to .r in the direction of "right-left." Accordingly, as the flux .r found the lesser resistance from Sº toward the left, so y would find the lesser resistance from Sº toward the right, if nothing had preceded. Actually the flux x has preceded the flux y immediately, and this fact, obviously, must be the cause which prevents the flux y from taking mainly the path toward the right of the diagram. To this causal relation we have to give expression in an assumption in the form of a general statement. We assume that any special kind of flux, say x, reduces not only the specific resistance, for x, of the neurons in which it occurs, but also the general resistance of these neurons; but we assume, in addition, that the reduction of the general resistance disappears under lack of function much more quickly than the reduction of the specific resistance.
Let us see what necessitates the latter half of this hypothesis: clearly, the admittance that any specific resistance of any neuron can be the result of function as well as of mere inheritance. If the reduction by function of the general resistance did not disappear more rapidly than the reduction of the specific resistance, no neuron could be said to have any specific resistance distinct from its general resistance, save directly by inheritance. On the other hand, our assumption implies that during a short time directly after the termination of a particular flux, the general resistance may be regarded as being as
( 163) much reduced as the specific resistance. It is plain that this is of the utmost importance for any immediate succession of two or more qualitatively different nerve processes.
Let us make the application. The flux x from the point S0,taking its path largely over the left side of the system of Figure 42, is immediately succeeded by the flux y, likewise from the point S0. If y had not been preceded by x, it would have found toward the right, over S2def, a resistance less, in proportion to the ratio (S+NG-G)/NS, than toward the left, over S2abc. But the general resistance of the left side having just been reduced, in consequence of the flux x, there may be for y no more resistance to the left than to the right. Suppose, then, for simplicity 's sake, that the flux y divides equally at the point S1 where the first branching occurs. The part passing to the right divides again at M2def. It divides about equally into N parts, since none of the neurons leading from M2def to the N motor points has a specific resistance for the flux y. The part passing from S1 to the left, divides at M2def also into N parts, --- but into unequal parts, since the neurons M2abc M1c, and M1c Mc, have a specific resistance for the flux y, all the others only the general resistance. If N is a rather high number, that is, if the peripheral branches are very numerous, the ratio of the flux reaching Mc to the flux reaching any other single motor point on either side of the diagram, must be approximately as S to G (The ratio S/G is the limit.)
Summarizing now the total motor influence of .r and y together, we reach the following conclusion. Of x but little flows into the motor points to the right. The main flow reaches the motor points on the left side, including Mc, but distributes itself equally over all these. Of y
( 164) a much larger (proportional to S/G) part flows into Mc than into any other motor point of the system. Therefore, the main motor response to the total stimulation x and y in this immediate succession occurs at Me. If the succession had been y and x, the main motor response, owing to the symmetrical relations of all the conditions concerned, would have occurred at Mf. We have thus demonstrated that the mere temporal relation of two stimuli (x and y) may result in a spatial determination (at Mc) of the motor response.
We wanted to explain the fact that a human being reacts to the word eat in one way, to the word tack in another way. In the diagram of Figure 4'2, which we have used for the explanation, we have supposed certain conductors to possess a specific resistance for one kind of flux, others a specific resistance for another kind. But that nature should by inheritance completely provide for such details of stimulation by such details of resistance distribution as those of this figure, is highly improbable. Our explanation is satisfactory only if we can show that a resistance distribution like that of Figure 42 may result from "experience" during the individual's life.
In the middle of Figure 42 we find a reflex arch whose central points are named S1c, and M1c.This is one representative of the whole class of reflex arches connecting the sensory points in the muscles and tendons of the speech organs with the motor points in the muscles of these same organs. These reflexes are undoubtedly of more importance for the talking instinct of the young of the human species than any other class of reflexes affecting the speech organs. In our figure the sensory point of the reflex arch has been marked by the word kinesthetic. This name is, of course, really too broad in its meaning, since it includes all sensory points found within any of the
( 165)
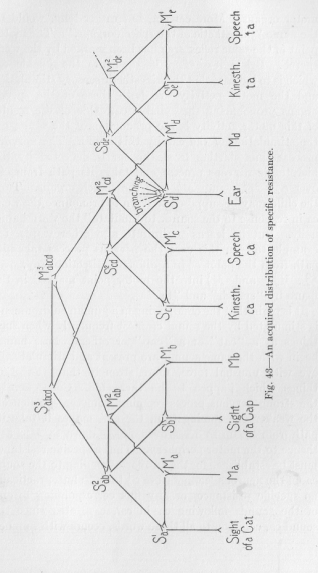
( 166) motor organs. More correct, but rather long, would be the name "kinesthetic of the speech organs." The motor point of the same reflex arch has been marked by the word speech. Below it we find iii the figure the syllable ca, which is to indicate that the kinesthetic sensory stimulation and the motor activity are both those of the speech organs pronouncing the syllable ca. One of the results or this motor activity is the production of the sound ca, the stimulation of the ear by this sound. The nervous process thus starting from the ear is deflected by the kinesthetic nervous process and takes its path from S1d over S2cd, M2cd, and M1c into the speech organs. Thus the child may be supposed to learn to pronounce the word ca in response to the sound, to "imitate" the word habitually. But he learns to imitate also other words, for example ta. In this case the nervous process takes its path from the point S1d in a different direction, say, over S2de, M2de and M1e into the motor point marked in the figure by "speech" and "ta."
It is plain that the child, living in a human environment, is stimulated by the sound ca most frequently when such things as a "cat" or a "cap" or others whose names begin with the same sound, are present and stimulate the eye with unusual force, more strongly than the other things which happen to be there too. Consequently, the strong nervous process coming from the eye deflects the weaker one coining from the ear, makes it take its path from S2cd up to S3abcd over M3abcd down to M2ab, and thence to the motor periphery. Thus a specific resistance must be established for the flux corresponding to the sound ca, in the neurons leading from S1d to M2ab; but practically no specific resistance for any flux corresponding to any of the sounds following ca in cat, cap, etc., since the sound ea, common to all these words, occurs with so much
(167) greater frequency. Thus the point S1d becomes, so to speak, a branching point of specific resistances, for ca to the left in the figure, for ta by a similar process to the right, and for many other sounds in other directions which are not explicitly drawn in Figure 43. This is exactly what we pre-supposed for the point S1 in Figure 4'2, with-out showing, at the time, how it could originate. The promise in this respect we have now made good. We still have to show, however, how in Figure 42 from the point M2abc, to a special motor point (Mc) a second kind of specific resistance can establish itself. Figure 43 answers our question. Whenever a cat is before the eye, the nervous process from the ear is drawn towards Ma, (in Figure 43); and this nervous process contains regularly the specific flux corresponding to the t-sound. Accordingly, the nervous path M2ab M1a Ma must have a very low specific resistance for this flux, compared with any other path branching off from M2ab. This explains the second presupposition embodied in Figure 42, for we had supposed, there, that one of the N branches from M2abc, to the periphery possessed a very low specific resistance for a second kind of flux. We were, therefore, entitled to the suppositions in Figure 42, and can rightly assert that we have demonstrated that the temporal order of qualitatively different stimuli may determine the motor point which responds.